東京湾広域アサリ浮遊幼生調査(アサリプロジェクト)
I. 概 要
- 1.はじめに
- 2.アサリプロジェクト実施までの道
- 2.1 どうやってアサリ浮遊幼生を分類するのか
- 2.2 いつ、どこで観測を行うのか
- 2.3 どうやってアサリの幼生を採集するか
- 2.4 観測方法の決定
1.はじめに
|
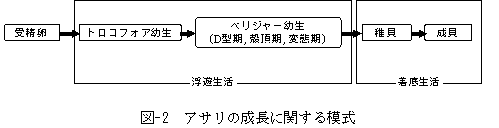
アサリRuditapes philippinarumは日本各地の干潟や浅場に生息し、潮干狩りなどのレクレーションの対象として、また食材として、日本人にとって最も馴染みのある二枚貝です。近年では、濾過食者としての高い海水浄化能力により、環境改善の面からも同種の重要性は高まっています。しかし,かつて15万トン前後あった漁獲量は、2000年には4万トン前後にまで落ち込んでいることから、アサリの現存量は全国的に激減していると考えられます。その主な原因として、東京湾では、アサリの主要な生息場所である干潟や浅場の埋め立てによる消失が挙げられています。また、生息場所の減少に伴う幼生供給のネットワークの衰退も、アサリをはじめとする干潟生物の減少要因として指摘されています(図I-1)。アサリは雌雄異体であり、放卵・放精による体外受精によって繁殖します。孵化した個体は、トロコフォア期およびベリジャー期幼生として2〜3週間水中を漂った後(一般に浮遊幼生あるいはプランクトン幼生と呼ばれています)、干潟や浅場などに着底して稚貝となり、成貝へと成長します(図I-2)。従って、東京湾においてアサリの資源を回復させるには、アサリ幼生の移流経路などを十分に把握した上で、生息場所の環境を保全、あるいは造成することが有効と考えられます。 |
|

|
近年、干潟や浅場の持つ水質浄化機能や、アサリを始めとする生物の生息場所、そして市民のレクレーションの場所としての機能が注目され、埋立地周辺に人工干潟を造成するなどの自然再生事業が計画されるようになりました(例えば、東京湾再生推進会議,2003)。幼生供給のネットワークを考慮することによって、これらの事業は、より効率的にアサリの生息場所を増やし、そしてアサリ資源の回復に繋がる可能性があります。また、アサリの現存量の増加に伴い、造成した場の水質浄化機能やレクレーション機能の向上が期待できます。海産の底生生物および付着生物の多くは浮遊幼生期を持つことから、浮遊幼生の移流経路に関する知見は、水産のみならず、環境保全および自然再生事業を行う海域を選定する際の基礎データとしても非常に重要と考えられます。そこで、国土技術政策総合研究所では東京湾におけるアサリの初期生態を解明することを目的として、運輸施設整備事業団と独立行政法人水産総合研究センター瀬戸内海区水産研究所と共同で、「東京湾広域アサリ浮遊幼生調査(通称アサリプロジェクト)」を2001年8月と10月に行いました。ここでは、アサリ幼生の調査を行うに当たり、どの様なことに気を付けて観測計画を立てたのか紹介します。なお、一連の報告は、2002年度および2003年度日本海洋学会春季大会で発表した内容と、当研究所で行われた平成14年度環境研修で用いた講義資料を基にまとめたものです。 |
2.アサリプロジェクト実施までの道
2.1 どうやってアサリ浮遊幼生を分類するのか
|
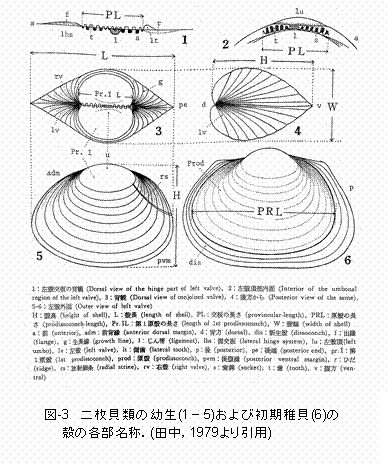
浮遊幼生の移流経路に関する情報を得るためには、東京湾全体を対象とした、時空間的に精密な調査が必要となります。しかし、幼生の大規模な調査はこれまで殆ど行われていませんでした。というのも、幼生の分類が非常に難しいのです。二枚貝類幼生は、左右の殻のずれを防ぐために、殻の交板上に交歯とじん帯があり、この構造(交装)は、殻の形状とともに、有効な分類形質となります(図I-3)。二枚貝類の殻の形成はベリジャー期幼生から始まります。ベリジャー期の幼生はその成長段階に応じてD型期(D-shaped
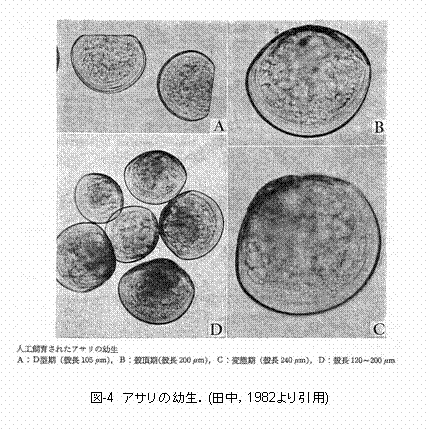
larva),殻頂期(umbo larva),そして変態期(fully grown larva)の3期に分けられます(図I-2)。D型幼生の殻は左右相称で、背縁は真直ぐか、あるいはその中央部がやや凹んでいます(図I-4)。D型期における交装は、一部を除き種で独得の特徴を表しません。また、殻の輪郭および大きさにも種間の差異が認めにくいと言われています。殻頂がふくらみ始めた殻頂初期以後の幼生は一般に殻頂期幼生と呼ばれ(図I-4)、交装は種特有の特徴を示します。さらに成長が進み、種固有の生活に移行できるまでに体制の整った幼生を変態期幼生と呼びます。 これまで、二枚貝の幼生の分類は形態や蝶番部分の構造の比較によって行われてき真下。しかし、脆い殻の取り扱いに注意が必要であることから、膨大な量のサンプルを処理するのに時間がかかりすぎてしまいます。また、蝶番が形成される前のD型幼生の分類が困難です。 |
|
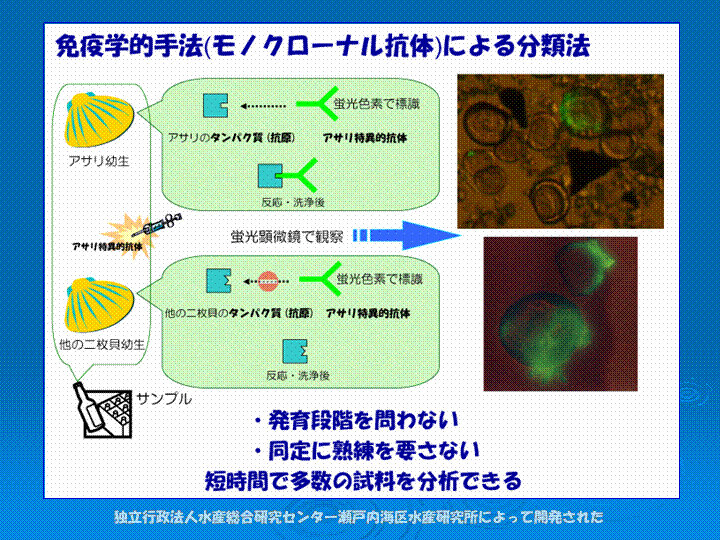
近年、免疫学分野の進歩によって、抗原抗体反応を用いてアサリ幼生を判別する方法が開発されました(図I-5)。この方法は光っている個体を数えるだけで良いので(図I-6)、短時間で多くのサンプルを処理することができ、大規模調査を行う上で非常に有効です。また、幼生の成長段階に関係なく、D型幼生も分類することができます。これによって、大規模調査の妨げとなる分類の問題をクリアすることができます。 |

2.2 いつ、どこで観測を行うのか
|
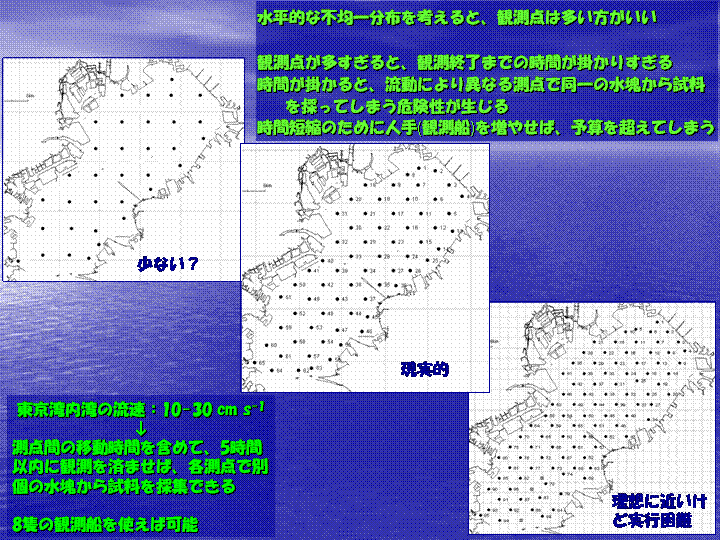
アサリ浮遊幼生をはじめとするプランクトンについてですが、プランクトンは水平的・鉛直的に不均一に分布します。また、プランクトンの分布には海況などの物理的要因、貧酸素水塊などの化学的要因、そして捕食などの生物的要因が作用すると考えられています。これはプランクトンの出現量を調べる上で非常に重要なことであり、これを考慮に入れてプランクトンの観測方法を考えなければなりません。 何処でアサリ浮遊幼生を採集するのか? 観測点の設定について考えてみます。東京湾では、アサリは盤洲や富津などに多くいます。しかし、他にも知られていない何処かにアサリが分布している可能性は十分に考えられます。従って、観測点は生息地の周りに重点的に置くのではなく、均一に設ける必要があります。また、プランクトンの水平的な不均一分布を考えると観測点は出来るだけ多くしたいところです。そこで測点を多くすると、今度は観測終了まで時間が掛かりすぎてしまい、水の動きによって、異なる測点で同じ水塊からサンプルを採ってしまう危険性が出てきます。時間をかけずに観測をするには観測船の数を増やせばいいのですが、今度は予算がオーバーしてしまいます。そこで、学術と予算の間でもっともバランスのとれた測点数が決まります(図I-7)。 図I-8に示した測点図では、およそ3.5 kmの間隔で65測点を設定しています。これらの測点でそれぞれ個別の水塊からアサリ浮遊幼生を採集するにはどうしたらいいか? 東京湾の流速が10〜30
cm s-1なので、間をとって20 cm s-1として考えると、5時間で3.6 km移動することになりますので、5時間以内で全ての観測を終えることができれば、各測点で異なる水塊からサンプルを採ることができると考えられます。これに、1測点あたりの観測にかかる時間と測点間の移動時間を含めて観測船の数を求めると、65測点を8隻の船を使って観測すれば5時間以内に観測を終える見込みがつきます。 |
|

|
次に、この観測を何時、何回行うか決めなければなりません。アサリの産卵は北の方では年一回、南の方では年2回行われます。東京湾では春先と夏〜秋の2回行われていると考えられています。本来であれば春と秋に観測を行うべきですが、予算決定などの事務手続き上の理由で、春期の観測は事実上不可能です。そこで、2回目の産卵期に的を絞って、8月と10月に観測を行うこととしました。何回行うかについてですが、アサリは、これまで多くの研究者によって行われてきた室内飼育実験の結果から、卵から孵化後2〜3週間で着底することが判っています。今回観測を行うのは8月と10月なので、高い水温で成長が早くなっていることも考慮して、2週間の内で数回行う必要があると考えられます。そこで本研究では、2001年8月2,6,10日と同年10月15,19,23日の日程で合計6回、観測を行うことにしました。これで観測をしたけどアサリ幼生が全く採れないという危険性を減らすことができます。 |
2.3 どうやってアサリの幼生を採集するか
|
アサリ浮遊幼生の採集方法としては、プランクトンネットを直接曳くことによる採集法と、ポンプで汲み上げた水をネットで濾過して集める方法が挙げられます。プランクトンネットによる採集では、ネットを鉛直方向に曳いてプランクトンを採集するのですが、これによりプランクトンの鉛直的な不均一分布の影響を受けにくいと言うメリットがあります。また短時間で採集することが出来ます。しかし、ネットの網目の目詰まりによって、濾水効率が低下してくる欠点があります。図I-9は目合100
µmと330 µmのネットを取り付けて、東京湾でプランクトンを採集している様子です。目合100 µmのネットは植物プランクトンなどで茶色くなっているのが判ります。アサリ浮遊幼生は小さいもので殻長100 µm程度ですので、目合50 µm程度の細かいネットで曳きたいところなのですが、あっという間に目詰まりしてしまうことが予想されます。 ポンプで汲み上げた水を濾過して採集する方法では、ポンプの排水速度を予め測れば、どれくらい濾過したのか判るという利点があります。また、目詰まりしても直ちに別のネットに交換することで容易に対処できます。また、100 µmのネットで濾過した海水を目合50 µmのネットで濾過することによって、小さなD型幼生の採集も可能となります。大量の水を汲み上げるのに時間がかかるという欠点もあります。 |
|

2.4 観測方法の決定
|
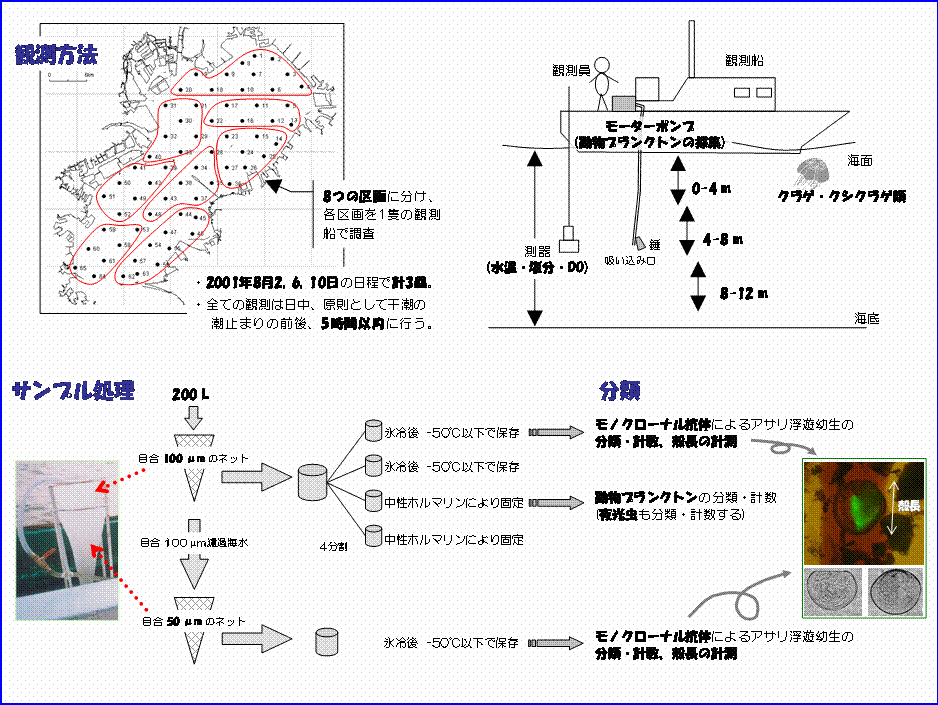
以上のことを考慮に入れて観測方法を決定しました(図I-10)。 東京湾は富津から観音崎を結んだ線を境にして北側を内湾、南側を外湾とし、区別して扱われることが多いですが、本研究で言う東京湾とは内湾を指します。観測は東京湾に約3.5 kmの間隔で設けた65測点で、2001年8月2,6,10日と同年10月15,19,23日の日程で計6回行うことにしました。全ての観測点を日中5時間以内に回れるように、65測点を8区画に分け、それぞれ1隻の調査船を用いて全区画同時に観測を行いました。アサリ浮遊幼生はモーターポンプによる採水法で採集しました。東京湾では底層は広い範囲で貧酸素状態となることから(風呂田,1988)、本研究では水深12 m以浅を採集の対象としました。採水は水深0〜4,4〜8,8〜12
mの3層で行い、観測点の水深に応じて採水層の数を変更しました。ポンプに繋がった吸水ホースの先端に錘を付け、吸い込み口を指定の採水層で繰り返し上下させながら200 Lの海水を汲み上げ、直ちに目合100 µmのハンドネットで濾過することによって幼生を捕集しました。濾過した海水をさらに目合50 µmのハンドネットで濾過することによって,より小さな幼生を捕集しました。目合100 µmのネットで得たサンプルについては分割器を用いて4分割し、2本は氷冷して持ち帰った後、-50℃以下で凍結保存、残り2本については中性ホルマリンが5%濃度となるように加えて固定保存しました。目合50 µmのネットで採集したサンプルについては1〜2分割し、氷冷して持ち帰った後、2本とも-50℃以下で凍結保存しました。凍結およびホルマリン固定した分割サンプルの内、それぞれ1本ずつを分析用、他を予備としました。 環境要因として、水温と塩分をアレック電子製STD(AST-500)によって測定し、両者の値から海水の密度(σt)を算出しました。溶存酸素量(DO)についてはYSI社製YSI-58型を用いて測定しました。観測中、肉食性の動物プランクトンである鉢クラゲ類のミズクラゲAurelia aurita、およびクシクラゲ類のカブトクラゲBolinopsis mikadoが確認された場合には、2×2 mの範囲内にいる個体数を船上から目視で計数しました。 本研究では、アサリ幼生の分類は、殻を有することで採集および保存時に破壊されにくく、また、個々の区別が容易なベリジャー期幼生を対象として行いました。アサリ幼生の分類には凍結保存した50 µmおよび100
µmネットサンプルを用いました。免疫学的手法(浜口,1999)によって処理したサンプル中の二枚貝類幼生を蛍光顕微鏡下で観察し、蛍光を発する個体をD型および殻頂期幼生に分けて計数しました。変態期の幼生については、殻頂期の幼生と形態的に大きな違いは無いことから殻頂期幼生として扱いました。免疫学的手法によるアサリ幼生の分類精度はおよそ95%と言われています(浜口,1999)。東京湾にはアサリの近縁種であるイヨスダレPaphia undulataが出現し(桑原,1990)、アサリ特異抗体は同種の幼生に対して反応します(浜口,未発表)。本研究では、殻頂期幼生については田中(1982)の分類データを基に形態による観察を併せて行い、蛍光を発するD型幼生で特異な個体についてはPCR法によって最終的な判別をしました。アサリと判定された幼生について最大100個体を無作為に選んで、その殻長を10 µmの単位まで計測しました。 アサリ幼生以外の動物プランクトンについては、16測点を対象として(図I-8参照)、ホルマリンで固定した100 µmネットサンプルを用いて実体顕微鏡下で種ごとに計数しました。渦鞭毛藻類の夜光虫Noctiluca scintillansは従属栄養生物であることから、本研究では動物プランクトンとして計数しました。 |
|
{kind=link}
次の章では、観測結果から明らかとなった東京湾におけるアサリ浮遊幼生の動態について紹介します。