東京湾広域アサリ浮遊幼生調査(アサリプロジェクト)
II. 夏季東京湾におけるアサリ浮遊幼生の動態
- 1.はじめに
- 2.海況
- 3. アサリ浮遊幼生の出現密度と殻長頻度分布
- 4.考察
- 4.1 ネットの定量性
- 4.2 幼生の殻長成長速度,発生日および浮遊期間の推定
- 4.3 幼生の発生場所
- 4.4 幼生の分布と環境要因との関係
- 5.おわりに
1.はじめに
|
「第I章 概要」において、研究の背景や、観測方法を決定するまでの過程について紹介しました。ここでは、2001年8月2,6,10日に行った観測結果を基に、アサリ幼生の発生場所や成長速度、浮遊期間について明らかにするとともに、幼生の時空間的な分布の変動要因について考察します。 |
2.海況
|
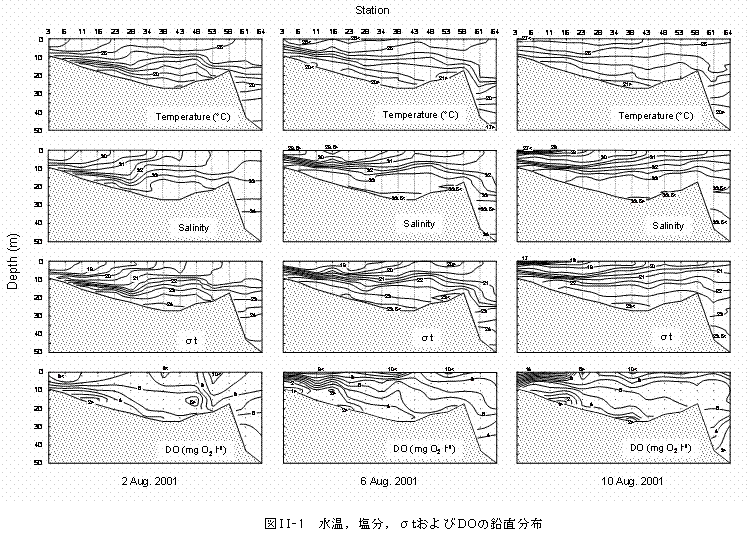
東京湾の湾軸に沿って、水温,塩分,σtおよびDOの鉛直分布を求めました(図II-1)。水温は17〜27℃、塩分は27〜34
psuの範囲となりました。密度躍層は8〜15 mの水深層に分布し、8月10日には湾奥の表層でも観察されました。DOは観測点3〜34間の海底付近では4 mg O2 ℓ-1以下であり、常に貧酸素の状態でした。8月6日には湾奥を中心に貧酸素水塊が水深5 m付近にまで分布しました。 水深0,4,8 mにおける海況の水平構造を図II-2に示します。水温は21〜28℃、塩分は17〜33
psuの範囲であり、湾奥では高温・低塩分、湾口では低温・高塩分の傾向が見られました。荒川河口域から多摩川河口域周辺の表層では、河川水の影響による塩分30 psu以下の水塊が観察されました。σtは塩分とほぼ同じ水平分布を示し、14〜22の範囲となりました。水深10 mにおけるDOは、湾奥の千葉側を中心とした広い範囲で4.5 mg O2 ℓ-1以下でした。 8月6日の海況は特徴的で、水温21〜25℃,塩分30〜33
psuの低温・高塩分の水塊が、千葉側に沿って湾口から湾奥まで分布しました(図II-2)。σtも同様で、盤州沖から市原にかけて顕著な密度フロントが観察されました。 |
|

3.アサリ浮遊幼生の出現密度と殻長頻度分布
|
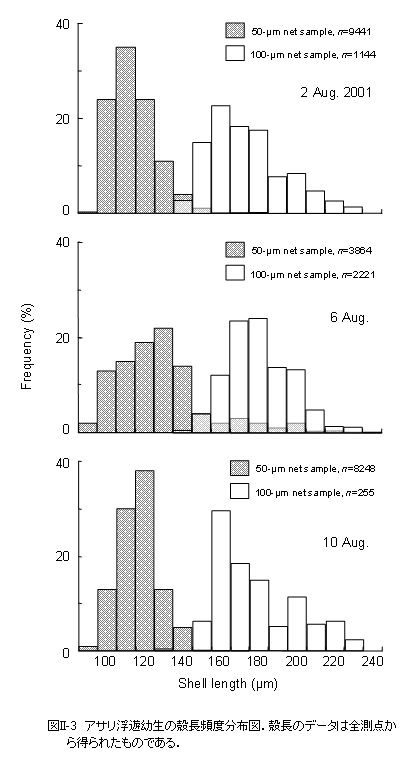
目合50および100
μmのネットによって殻長90〜240 μmの幼生が採集されました(図II-3)。D型幼生の殻長は90〜130 μm、殻頂期幼生の殻長は130〜240
μmでした。殻長組成は2〜3峰型を示し、ピークは8月2日には殻長110および160
μmに、8月6日には殻長130および170〜180
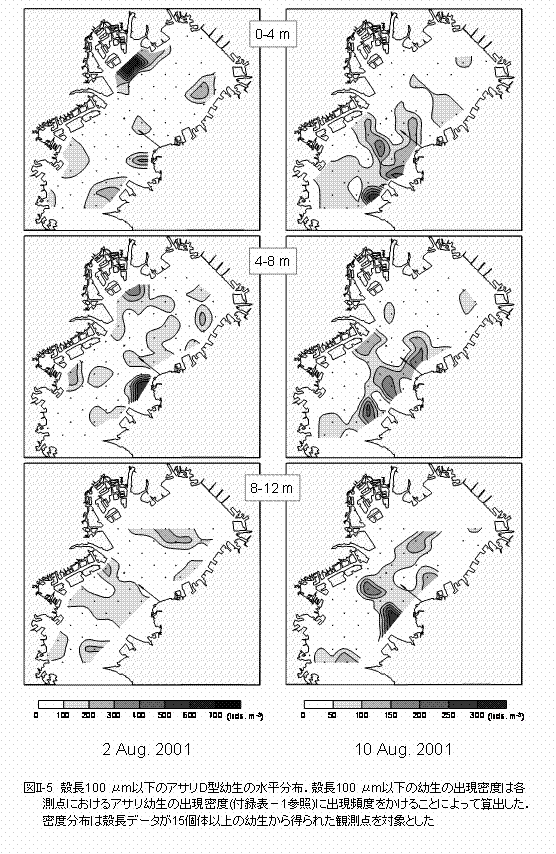
μmに現れました。8月10日の殻長のピークは120,160,そして200 μmでした。 幼生はほぼすべての観測点から採集されました(図II-4)。8月2日と10日にはD型幼生が多量に出現し、最大出現密度はそれぞれ2510個体m-3および1690個体m-3に達しました。8月6日には殻頂期幼生が非常に多く出現し、最大出現密度は2725個体m-3に達しました。 D型幼生が多く出現した8月2日、および8月10日の出現密度と殻長頻度のデータを用いて殻長100 μm以下のD型幼生の出現密度を算出し、その水平分布を求めました(図II-5)。殻長100
μm以下のD型幼生は、三枚洲〜羽田および盤洲周辺の海域に500個体m-3以上の高い出現密度で分布しました。また、横浜や市原、そして富津周辺の海域にも200個体m-3以上の出現密度で小型の幼生は分布しました。 |
|

4.考察
|
殻長140 μmのアサリ幼生は100 μmネットサンプルからだけではなく、50 μmネットサンプルからも多く観察されたことから(図II-3)、殻長140
μm以下の幼生は目合100 μmのネット、すなわち殻長に対して70%の目合のネットでは正確に定量できません。アサリの幼生は、初期のD型幼生で殻長100 μm程度であることから(鳥羽,1987)、目合50 μmのネットを使うことによって、D型期以降の幼生をほぼ逃さずに採集できると考えられます。従って,本研究で得られたアサリD型および殻頂期幼生に関するデータは、東京湾におけるアサリ浮遊幼生の殻長組成や出現密度を反映していると思われます。 |
4.2 幼生の殻長成長速度,発生日および浮遊期間の推定
|
今回の観測で採集された幼生の最大殻長は240 μmであり、殻長210
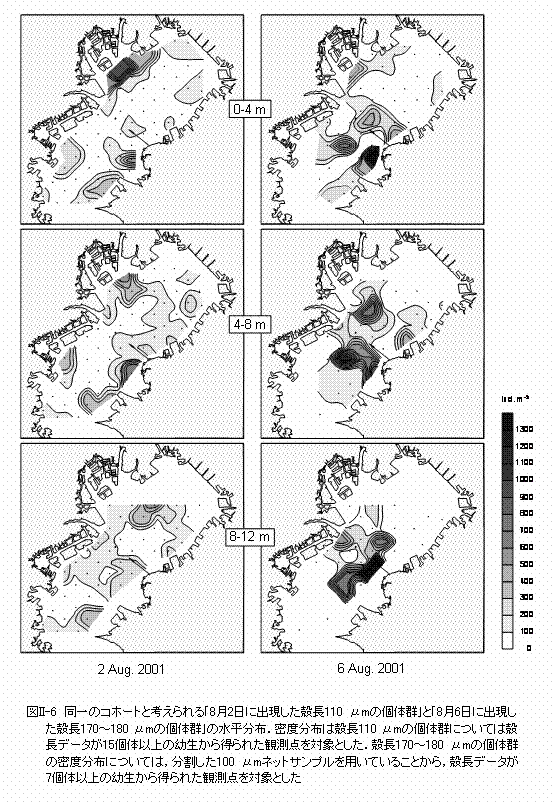
μm以上の幼生の出現頻度は低下しました(図II-3)。室内飼育実験におけるアサリ幼生の着底時の殻長はおよそ185〜230 μmであることから(鳥羽1987参照)、東京湾においても、殻長が210 μm前後に達した幼生は着底生活に移行すると思われます。 幼生は、8月2日と10日にはD型期、8月6日には殻頂期というように、どちらか一方の成長段階が優占して現れたことから(図II-4)、幼生の出現密度の時間的な変動は主に一連の成長過程によるものと思われます。すなわち、8月2日に最も個体数の多かった殻長110 μmのD型幼生は、8月6日には最も優占した殻長170〜180 μmの殻頂期幼生に成長したと考えられます(図II-3)。この同一と考えられる個体群の出現密度の変化から、個体群密度の減少率は4日間でおよそ13%と推定されました。また、殻長の変化から、夏季の東京湾における幼生の殻長成長速度は、1日当たり15〜18
μmに達することが明らかとなりました。この個体群は8月10日には殻長210
µm以上にまで成長することが可能であることから、この時までには大部分が浮遊生活から着底生活に移行したと考えられます。 殻長成長速度が判れば、幼生が生まれた日を逆算することができます。アサリの卵は水温20℃の条件下では、受精後2日程度で殻長100 μm前後のD型幼生となることから (鳥羽,1987)、8月2日に優占した殻長100〜120 μmの幼生は、D型幼生となってから0〜2日、生まれてから2〜4日経過しています。従って、これらの幼生が生まれたのは7月30日前後と推定できます。同様に、8月10日に優占した殻長110〜120 μmの幼生については、8月6日前後に生まれたと考えられます。8月2日に優占した殻長100〜120 μmの幼生の多くは8月10日までには着底生活に移行したと考えられることから、夏季の東京湾におけるアサリ幼生の浮遊期間は10日程度であることが明らかとなりました。 では、これらの幼生はどこで生まれたのでしょうか? |
4.3 幼生の発生場所
|
殻長100 μm以下の幼生が多く分布した羽田〜三枚洲や盤洲周辺の海域は、アサリ成貝の生息域とほぼ一致します(図I-8参照)。殻長100 μm前後のD型幼生は卵として放出されてから2日程度であることから(鳥羽,1987)、生まれた場所からそれほど分散していないと思われます。従って、羽田〜三枚洲と盤洲周辺の海域に加えて、小型幼生が多数分布した横浜や市原および富津周辺の海域でアサリの幼生は発生したと考えることができます。 盤洲や富津では干潟や浅場が自然に近い状態で残っており、アサリの成貝の資源量も大きいことから(鳥羽,2002)、多量の幼生が発生していると考えられます。三枚洲〜羽田周辺の海域については、多摩川河口域(桑原,1990) や三枚洲周辺(風呂田,1997) にアサリが生息している。この海域におけるアサリの資源量に関する定量的な研究は極めて少ないですが、D型幼生の出現密度は盤洲や富津周辺の海域に匹敵することから、三枚洲〜羽田周辺海域のアサリの産卵個体群は、東京湾における幼生の供給に大きく寄与していると思われます。横浜および市原周辺の海域は港湾域であり、主に鉛直護岸で囲まれています。市原周辺の地先の海底には砂が堆積しており、そこにはアサリが生息している(鳥羽,私信)。また、横浜周辺の海域でもアサリが生息していることから(桑原,1990)、港湾域のような水深のある場所でも、砂の堆積などのアサリの生息条件が整えば、幼生の供給場所として機能することが示唆されました。 三番瀬は盤洲と並びアサリの資源量が多いことから(鳥羽,2002)、周辺の海域からは多くの幼生が採捕されることが予想されましたが、その出現量は少ない結果となりました。三番瀬では、青潮や江戸川からの淡水の大量出水によるアサリの大量斃死が、夏から秋にかけてしばしば発生します(柿野,1992参照)。突発的な生息環境の悪化によるアサリの産卵個体群の激減が、三番瀬周辺海域における幼生の発生量の減少に繋がっている可能性があります。金沢湾には人工海浜が造成されており、そこに多くのアサリが生息しています。また、金沢湾に隣接する平潟湾にも非常に多くのアサリが生息しています(新保ら,1999)。金沢湾では4〜5月の連休を中心に、多くの観光客によって潮干狩りが行われ、殻長20 mm以上の個体群に対して非常に高い漁獲圧が掛かかります(工藤,2000)。アサリは殻長20 mm程で成熟することから(鳥羽・深山,1994)、過度の漁獲による成熟個体の激減が、この海域における幼生の発生量を制限していると思われます。平潟湾では潮干狩りが行われていないことから、アサリの成熟個体の出現量は安定しています(越川ら,1999)。平潟湾は金沢湾内の野島海岸への幼生の供給源として考えられていますが、さらに沖合の東京湾への幼生供給の役割については、幼生の沖合への輸送に寄与する流れなどのプロセスを含めて今後さらに検討が必要でしょう。 |
4.4 幼生の分布と環境要因との関係
|
浮遊生物は遊泳力が小さいことから、その分布は水塊の動きなどに大きく影響されます。また、浮遊生物の卵や幼生が底層の貧酸素水塊に輸送されることによって死亡し、結果として出現密度の減少に繋がることもあります。餌量や被捕食も浮遊生物の出現密度の変動要因として挙げられます。では、東京湾のアサリ幼生の分布にはどの様な要因が作用していたのでしょうか? |
|
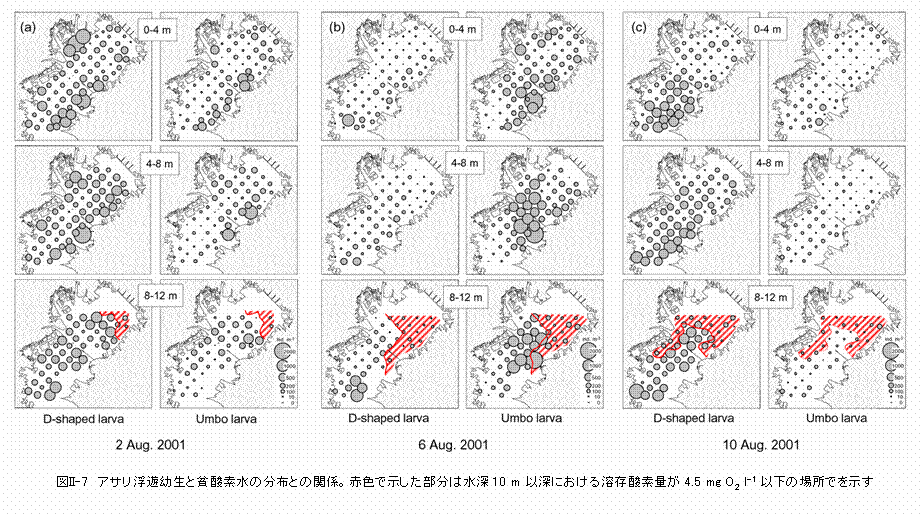
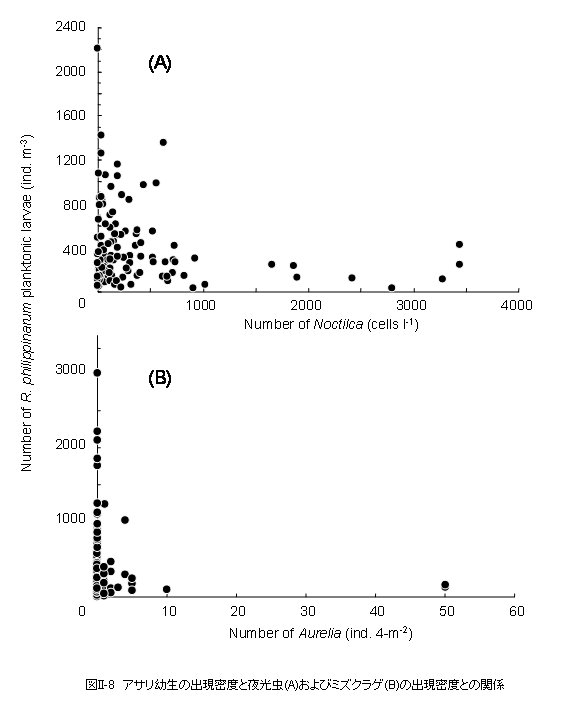
アサリ浮遊幼生は東京湾の全域から採集されたことから、浮遊している間に広範囲に拡散していることが明らかとなりました(図II-4)。そこで、同一の個体群である「8月2日に出現した殻長110μmの個体群」と、「8月6日に出現した殻長170〜180μmの個体群」の水平分布を比較してみます(図II-6)。三枚洲〜羽田や盤洲などで発生した幼生は、8月6日には顕著な密度フロント(図II-2参照)が存在する湾の中央部に移動していることが判ります。従って、今回観察されたアサリ幼生の出現密度の変動には、移流による個体群の湾外への流出はそれほど作用していないと思われます。8月6日に観察された密度フロントは強い北風によって引き起こされたものであり、夏季の東京湾では比較的よく観察されます。フロントに沿った海面上にはゴミやクラゲなどが集積することが知られており、アサリ幼生の水平分布には物理的な収束機構が強く作用することが示唆されました。 貧酸素水塊の影響はどうでしょうか? 観測期間中、東京湾の水深10 mにおけるDOは湾奥を中心として広い範囲に渡って貧酸素の状態にあり、この範囲内にある観測点では、水深8〜12 m層における幼生の出現密度は他の観測点よりも低い傾向が見られました(図II-7)。しかし、これらの観測点では、貧酸素の状態にない水深4 m以浅の層でも幼生の出現密度は低いことから、この時期に見られた程度の貧酸素水塊(4 mg O2 ℓ-1以下)の幼生の分布に対する影響は不明です。 次は捕食者の影響を見てみましょう。今回の調査では、肉食性の動物プランクトンとしては、夜光虫,カイアシ類のOncaea media,ヤムシ類のSagitta crassa,そしてミズクラゲなどが観察されました。中でも夜光虫とミズクラゲは多く出現し、最大出現密度はそれぞれ3500細胞 ℓ-1および50個体4-m-2に達しました。夜光虫のアサリ幼生に対する捕食速度については知見が得られておらず、今後、室内実験などにより明らかにしていく必要があります。ミズクラゲの出現密度については、本研究では表層の出現数に関する情報しか得られていないため、ミズクラゲ個体群のアサリ幼生に対する捕食圧を正確に見積もることはできませんが、夜光虫およびミズクラゲが大量に出現した観測点ではアサリ浮遊幼生は少ない傾向が見られました(図II-8)。夜光虫、ミズクラゲとも高密度の個体群がパッチ状に分布することから、両種による捕食圧は幼生の出現密度に対して局所的に作用している可能性があります。 |
|
{kind=link}
{kind=link}
{kind=link}
5.おわりに
|
8月の調査によって、自然の干潟や浅場以外に港湾域のような深場もアサリ幼生の供給源として機能していることが明らかとなりました。港湾域は河口域ほど環境変動が大きくない上に、アサリの成熟個体群に対して潮干狩りなどによる漁獲圧が掛からないことから、東京湾における安定した幼生の供給源であるのかもしれません。近年、干潟や浅場の持つ浄化機能や生物の生息場としての役割が注目され、埋立地周辺に人工干潟を造成するなどの自然再生事業が計画されています。これまで生物の生息場所としての内湾港湾域の重要性は殆ど認識されていませんでしたが、事業などを行う際には、同海域の生物分布に関する詳細な調査を行う必要があるでしょう。 |