�����p�L��A�T�����V�c�������i�A�T���v���W�F�N�g�j
III.�@�H�G�����p�ɂ�����A�T�����V�c���̓���
- 1�D�͂��߂�
- 2�D�C��
- 3. �A�T�����V�c���̏o�����x�Ɗk���p�x���z
- 4�D�l�@
- 4.1�@�A�T���c���̊k���������x�C�������C����ѕ��V���Ԃ̐���
- 4.2�@�c���̔����ꏊ
- 4.3�@�c���̕��z�Ɗ��v���Ƃ̊W
- 5�D������
�P�D�͂��߂�
|
8���̊ϑ��ɂ���āA�@���R���邢�͐l�H�̊������ꂾ���ł͂Ȃ��A�`�p���̂悤�Ȑ��[�̂���ꏊ���A�T���c���̋����ꏊ�Ƃ��ċ@�\���Ă��邱�ƁA�A�c���̊k���������x��1��������15�`18
��m�ɒB���A��10���Ԃ̕��V�������߂�������A�k��210 ��m���x�Œ��ꐶ���Ɉڍs���邱�ƁA�B�A�T���c���̕��z�ɂ͖k���ɂ���Ĉ����N�����ꂽ���x�t�����g�ɂ������@�\��A�~�Y�N���Q����і�����ɂ��ߐH����p���Ă��邱�ƁA�Ȃǂ���������܂���(��II��4�D4�Q��)�B�����p�̊C���͋G�߂ɂ���đ傫���قȂ�A10���̐�����8�������啝�ɒቺ���܂��B�܂��A8���͓�Ε�����z����̂ɑ��āA10���ɂ͖k�Ε�����z���܂�(��2�`�p���ǁC1974�Q��)�B�]���āA10���ɂ�����A�T���c���̐�����c�ߒ��́A8���Ƃ͑傫���قȂ邱�Ƃ��\�z����܂��B����ɁA8���̊ϑ��ł͊m�F�ł��Ȃ������c���̋����������݂���\�����l�����܂��B�����ł́A�u�����p�L��A�T�����V�c�������v��10���̊ϑ����ʂ���ɁA�A�T���c���̔����ꏊ������x�A���V���Ԃɂ��Ė��炩�ɂ���ƂƂ��ɁA�c���̎���ԓI�ȕ��z�̕ϓ��v���ɂ��čl�@���܂��B |
�Q�D�C��
|
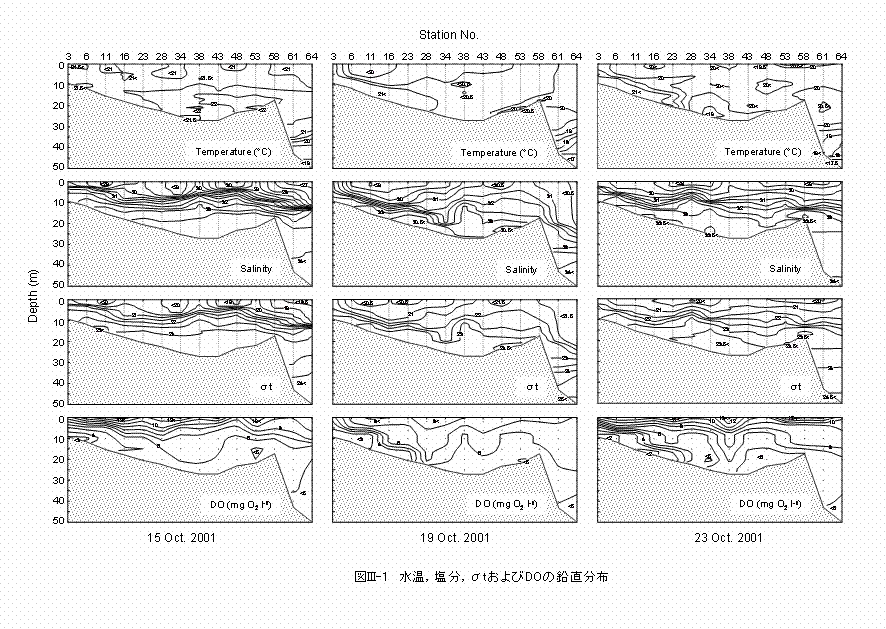
�����p�̘p���ɉ����āA�����C�����C��t�����DO�̉������z�����߂܂���(�}III-1)�B������17�`22���͈̔͂ł����B���[0�`25 m�w�ɂ����鐅����21���O��ł���A�����I�ɂقڈ�l�ł����B������27�`34�͈̔͂ł���A���[5�`15 m�ɖ��w���ώ@����܂����B��t��18�`25�͈̔͂ł���A�����Ƃقړ��l�̉������z�������܂����B���x���w�͐��[5�`15 m�ɕ��z���܂����B10��17������18���ɂ����đ䕗21�����֓��t�߂�ʉ߂������ʁA���x���w��10��19���ɂ͑傫������܂������A10��23���ɂ͐��[8 m�O��Ɍ`������܂����BDO�ɂ��ẮA4 mg O2 ℓ-1�ȉ��̕n�_�f����́A���������ɂ͎��炸�A10��19�������23���ɂ͊ϑ��_3�`23�ɂ����ĊC��t�߂ɕn�_�f�����z���܂����B ���[0�C4�C8 m�ɂ�����C���̐����\�����}III-2�Ɏ����܂��B������21���O��ł���A�����I�ɂ��قڈ�l�ƂȂ�܂����B������15�`32�͈̔͂ł���A�k�Ε����������Ƃ��̓����ł���p���ɉ��������l���̕��z�������܂����B�ቖ�����͉͐쐅�̉e������p�������̐��[0 m�Ɏ�ɕ��z���܂����B10��15���̐��[0 �����4 m�ɂ́A10��9���Ɋ֓��t�߂�ʉ߂����䕗20���ɂ�鍋�J�̉e���Ǝv����ቖ�������A�����s����_�ސ쌧���ɉ������L���͈͂Ŋώ@����܂����B��t�͉����Ƃقړ����������z�������A9�`23�͈̔͂ł����B�͐쐅�̉e������p���̐��[0 m�A����ё䕗�̍��J�ɂ��Ǝv����10��15���̐��[0�����4 m�������āA��t�̐����I�ȕϓ��̌��z�͊ɂ₩�ł����B���[10 m�ɂ�����DO�́A�p����4.5
mg O2 ℓ-1�ȉ��̕n�_�f��ԂƂȂ��Ă�����̂́A���͈̔͂�8���Ɣ�r���Ă��悻�������x�ɏk�����܂����B |
|

�R�D�A�T�����V�c���̏o�����x�Ɗk���p�x���z
|
8���̊ϑ��ɑ����A��ʂ̃A�T���c�����̏W���邱�Ƃ��ł��܂����B�ڍ�50�����100
��m�̃l�b�g�ɂ���Ċk��90�`230 ��m�̗c�����̏W����܂���(�}III-3)�BD�^�c���̊k����90�`130 ��m�A�k�����c���̊k����130�`230
��m�ł����B�k���g����1�`2��^�������A�s�[�N��10��15���ɂ͊k��120�`140
��m�ɁA10��19���ɂ͊k��110�����160�`170
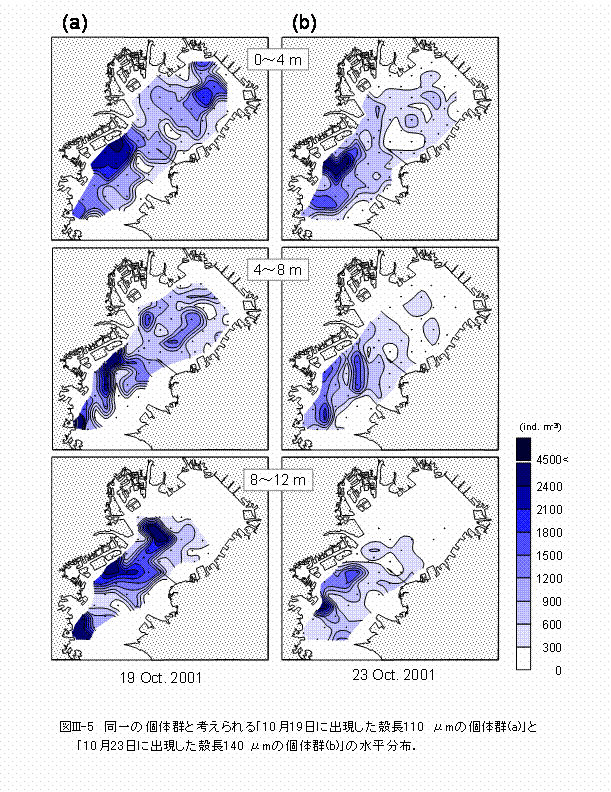
��m�Ɍ���܂����B10��23���̊k���̃s�[�N��140 ��m�ł����B �c���͂قڂ��ׂĂ̊ϑ��_����̏W����܂���(�}III-4)�B10��19���ɂ�D�^�c�������ʂɏo�����A�ő�o�����x��5390��m-3�ɒB���܂����B10��23���ɂ͊k�����c�������ɑ����o�����A�ő�o�����x��2670��m-3�ɒB���܂����B D�^�c���������o������10��19���̏o�����x�Ɗk���p�x�̃f�[�^��p���āA�k��110 ��m�ȉ���D�^�c���̏o�����x���Z�o���A���̐������z�����߂܂���(�}III-5a)�B�k��110
��m��D�^�c���́A�O�Ԑ��C�O���F�`�H�c�C�s���C���`���l�C�����ċ���p���ӂ̊C��ɁA1000�`4800��m-3�̔��ɍ������x�ŕ��z���܂����B |
|

�S�D�l�@
4�D1�@�A�T���c���̊k���������x�C�������C����ѕ��V���Ԃ̐���
|
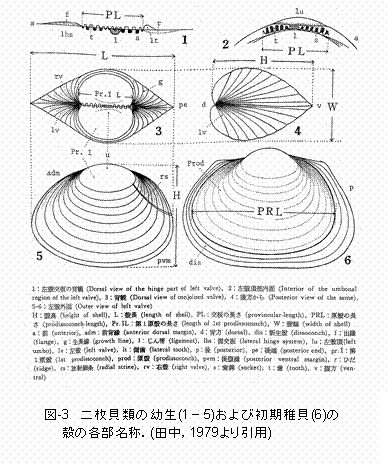
�u�����p�L��A�T�����V�c�������v�ł͖ڍ�100 ��m�̃l�b�g�ƕ����āA�ڍ�50 ��m�̃l�b�g���g�����Ƃɂ���āAD�^�����܂߂��A�T���c���̐��m�Ȓ�ʂ��\�ƂȂ�܂���(��II��4�D1�Q��)�B�{�����œ���ꂽ�f�[�^�͓����p�ɂ�����A�T���c���̏o���f���Ă���ƍl�����܂��B�A�T���c����10��19���ɂ�D�^���A10��23���ɂ͊k�����Ƃ����悤�ɁA�ǂ��炩����̐����i�K�̌̌Q���D�肵�Č��ꂽ���Ƃ���(�}III-4)�A�c���̏o�����x�̎��ԓI�ȕϓ��͎�Ɉ�A�̐����ߒ��ɂ����̂Ɛ��@����܂��B���Ȃ킿�A10��19���ɍł��̐��̑��������k��110 ��m��D�^�c���́A10��23���ɂ͍ł��D�肵���k��140 ��m�̊k�����c���ɐ��������Ǝv���܂�(�}III-3�Q��)�B���̓���ł���̌Q�̏o�����x�̕ω�����A�A�T���c���̌̌Q���x�̌�������4���Ԃł��悻24���Ɛ��肳��܂����B���̒l��8���ɂ����錸�����̂��悻2�{�ł�(�\III-1)�B���l�ɁA�Ԃɑ䕗���ʉ߂���10��15������19���ɂ����ẴA�T���c���̌̌Q���x�̌������A����̌̌Q�ƍl������u10��15���ɗD�肵���k��120�`140 ��m�̌̌Q�v�Ɓu10��19���ɗD�肵���k��150�`180 ��m�̌̌Q�v�̏o�����x�̕ω����狁�߂�����(�}III-3�Q��)�A4���Ԃł��悻72���ɒB���邱�Ƃ����炩�ƂȂ�܂����B�܂��A�����̌̌Q�̊k���̌o���ω�����A�A�T���c���̊k���������x��1��������8�`9
��m�Ɛ��肳��A8���̊k���������x�����啝�ɒቺ���Ă��邱�Ƃ����炩�ƂȂ�܂���(�\III-1)�B |

|
�k���������x����c���̔���������ѕ��V���Ԃ𐄒肷�邱�Ƃ��ł��܂��B����20���̏������ł́A�A�T���̗��͕��o���ꂽ��2���Ŋk��100 ��m���x��D�^�c���ɐ������邱�Ƃ���(���H�C1987)�A10��19���ɗD�肵���k��110 ��m�̗c����D�^�c���ƂȂ��Ă���1�`2���A���Ƃ��ĕ��o����Ă���3�`4���o�߂��Ă���ƍl�����܂��B�]���āA���̌̌Q�����Ƃ��ĕ��o���ꂽ�̂�10��15���`16�����Ɛ��肳��܂��B 10���̊ϑ��ō̏W���ꂽ�c���̍ő�k����230 ��m�ł���A�k��210
��m�ȏ�̗c���̏o���p�x�͒ቺ���܂���(�}III-3)�B8���̊ϑ��ł��k��210 ��m�ȏ�̗c���̏o���p�x�͒ቺ�������Ƃ���A�����p�ɂ�����A�T���c���͊k��210 ��m�O��ŕ��V�������璅�ꐶ���Ɉڍs���Ă���ƍl�����܂��B������������ɂ�����A�T���c���̒��ꎞ�̊k���͂��悻185�`230
��m�ł���(���H�C1987�Q��)�A�{�������ʂ��x�����Ă��܂��B10��23���ɗD�肵���k��140 ��m�̗c�����A�k���������x8�`9
��m d-1�Ŋk��210 ��m�ɒB����̂͂��悻8����A���Ȃ킿10��31���O��ł��B���̌̌Q�́u10��19���ɗD�肵���k��110 ��m�̌̌Q�v�Ɠ���ł���A10��15���`16���ɗ��Ƃ��ĕ��o���ꂽ���Ƃ���(��L�Q��)�A10���̓����p�ɂ�����A�T���c���̕��V���Ԃ͂��悻15���ƍl�����܂��B 8���̃A�T���c���̕��V���Ԃ͂��悻10���ł��邱�Ƃ���A10���ɂ�����c���̕��V���Ԃ�8���Ɣ�ׂ�5���Ԃقǒ����Ȃ邱�Ƃ����炩�ƂȂ�܂���(�\III-1)�B���ꐶ���Ɉڍs����k���T�C�Y��8�������10���Ƃ���210
��m�ł���A�����̊ԂɈႢ�͂Ȃ����Ƃ���A10���ɂ�����A�T���c���̕��V���Ԃ̉����́A�k���������x�̒ቺ�ɂ����̂Ǝv���܂��B�A�T���c���͉a�Ƃ���10 ��m�ȉ��̏��^�̐A���v�����N�g����ߐH����ƍl�����Ă��܂�(���X�C2001)�B�ϑ����s�������Ԃ̓����p�ɂ����鏬�^�̐A���v�����N�g���̏o���ʂɂ��Ă͕s���ł��B�������A�{�����œ���ꂽ�c���̊k���������x�́A���H(1992)������������瓾���k���������x(����21���ɂ�����4.4
��m d-1)�������ɍ������Ƃ���A10���̓����p�ɂ�����A�T���c���͉a�ʂɂ�鐬���̐������Ă��Ȃ��Ǝv���܂��B10���ɂ�����A�T���c���̊k���������x�̒ቺ�́A�����������������Ƃɂ��c���̊������ቺ�������߂ł���ƍl�����܂��B |
4�D2�@�c���̔����ꏊ
|
�k��110 ��m�̗c�����������z�����O�Ԑ��C�O���F�`�H�c�C�����ċ���p���ӂ̊C��́A�A�T���̐�����Ƃقڈ�v���܂�(�}I-8�Q��)�B�k��110
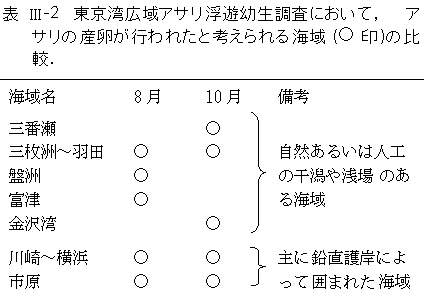
��m�O���D�^�c���͗��Ƃ��ĕ��o����Ă���3�����x�ł��邱�Ƃ���A���܂ꂽ�ꏊ���炻��قǕ��U���Ă��Ȃ��Ǝv���܂��B�]���āA�O�Ԑ��C�O���F�`�H�c�C�����ċ���p���ӂ̊C��ɉ����āA���^�c���������̏W���ꂽ���`���l����юs�����ӂ̊C��ŁA�A�T���̗c���͎�ɔ��������ƍl�����܂��B 8���̊ϑ��ɑ����A�A�T���̎Y���͍L�͈͂łقړ����ɋN���Ă��邱�Ƃ����炩�ƂȂ�܂���(�\III-2)�B�A�T���̎Y���͑䕗�Ȃǂɂ�鐶���ꏊ�̝����Ȃǂɂ���ėU������邱�Ƃ��m���Ă��܂�(������C2001)�B�ϑ����Ԓ��A�䕗21����10��17������18���ɂ����Ċ֓��t�߂�ʉ߂��܂����B�������A�A�T���c�������܂ꂽ�ƍl������10��15���O��ɂ́A�䕗21���͑�p�t�߂ɂ��������Ƃ���A�䕗�ɂ�鐶���ꏊ�̝������A����ώ@���ꂽ�A�T���̓����Y����U�������Ƃ͎v���܂���B�A�T���̓����Y���̗U���v���ɂ��ẮA���コ��ɒ������K�v�ł��B |
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
8���̊ϑ��ł͔��ɑ����̏��^�c�����̏W���ꂽ�ՏF����ѕx�Î��ӂ̊C�悩��́A10���̊ϑ��ł͏��^�c���͖w�Ǎ̏W����܂���ł���(�}III-5a)�B�ՏF��x�Âɂ͎��R�̊������ꂪ�c����Ă���A�����̊C��ɂ�����A�T���̌����ʂ͔��ɑ傫���ƌ����܂�(���H�C2002)�B�����āA�ϑ����s���������ɁA�ՏF��x�Âɂ�����A�T���̑�ʝˎ��Ɋւ���͖������Ƃ���A�����̊C��ŏ��^�c�������Ȃ������̂͐e�L�̌����Ȃǂɂ����̂ł͂Ȃ��A��K�͂ȎY�����s���Ȃ��������Ƃɂ��ƍl�����܂��B�A�T���̑�K�͎Y���̃^�C�~���O����їU���v���́A�C�悲�ƂɈقȂ邱�Ƃ���������܂��B �O�Ԑ������p�ɂ͎��R���邢�͐l�H�̊������ꂪ����A�����ɂ͑����̃A�T�����������Ă��܂�(�V�ۂ�C1999�G���H�C2002)�B�������A8���̊ϑ��ł́A���ӂ̊C�悩��͏��^�c���͖w�Ǎ̏W����܂���ł���(�\III-2)�B���̌����Ƃ��āA�O�Ԑ��ł͐���]�ː삩��̒W���̑�ʏo���A����p�ł͒������A�ɂ��A�T���e�L�̌�������������܂���(��II��4�D3�Q��)�B�O�Ԑ������p�ł̓A�T���̎�c�������s���Ă��Ȃ����Ƃ���A�e�L�̐l�דI�ȑ����͋N����Ȃ��͂��ł��B�A�T���̒t�L�́A�ċG�̍��������������ł͋}���ɐ������邱�Ƃ��\�ł��邱�Ƃ���(�`��E���H�C1990)�A�ċG�ɐ����c�����t�L���������A10���܂łɂ͎O�Ԑ�����ы���p�ɂ�����e�L�̌̌Q�ɉ��������ƍl�����܂��B �O���F�`�H�c�C�s���C�����Đ��`���l���ӂ̊C�悩��́A8���̊ϑ��Ɉ��������A���ɑ����̏��^�c�����̏W����܂���(�\III-2)�B�O���F�`�H�c���ӂ̊C��ɂ����R���邢�͐l�H�̊������ꂪ���݂��A�A�T���͑�����͌���(�K���C1990)��O���F����(���C�c�C1997)�ɐ������Ă��܂��B�s������`���l���ӂ͍`�p���ł���A���͎͂�ɉ�����݂ɂ���Ĉ͂܂�Ă��܂��B�s�����ӂ̒n��̊C��ɂ͍����͐ς��Ă���A�����ɂ̓A�T�����������Ă��܂�(���H�C���M)�B�܂��A���l���ӂ̊C��ɂ��A�T���̐������m�F����Ă��܂�(�K���C1990)�B�����̊C��ɂ�����A�T���̌����ʂɊւ����ʓI�Ȍ����͋ɂ߂ď��Ȃ��ł����AD�^�c���̏o�����x�͎O�Ԑ������p���ӂ̊C��ɕC�G���邱�Ƃ���A�O���F�`�H�c�C�s���C�����Đ��`���l���ӊC��̃A�T���̎Y���̌Q�́A�����p�ɂ�����c���̋����ɑ傫����^���Ă���Ǝv���܂��B�܂��A���R����ѐl�H�̊������ꂾ���ł͂Ȃ��A�`�p���̂悤�ɐ��[�̂���ꏊ���A���̑͐ςȂǂ̃A�T���̐��������������A�c���̋����ꏊ�Ƃ��ċ@�\���邱�Ƃ���������܂����B |
4�D3�@�c���̕��z�Ɗ��v���Ƃ̊W
|
8���̊ϑ��Ɉ��������A10���̊ϑ��ɂ����Ă��A�T���c���͓����p�̑S�悩��̏W����܂���(�}III-4)�B�A�T���c���͕��V���Ă���ԂɍL�͈͂Ɋg�U���Ă��邱�Ƃ���A�c���͔��������ꏊ�Ƃ͈قȂ�ꏊ�ɒ��ꂵ�Ă���\��������܂��B����̌̌Q�ł���A�u10��19���ɏo�������k��110 ��m�̌̌Q�v�Ɓu10��23���ɏo�������k��140 ��m�̌̌Q�v�̐������z���r���Ă݂܂�(�}III-5)�B�O�Ԑ��C�O���F�`�H�c�C���`���l�C�����ċ���p�ȂǂŔ��������c���́A���[0�`4 m�����4�`8 m�w�ɂ�����̌Q�ɂ��Ă͘p���ցA���[8�`12 m�w�ɂ�����̌Q�͘p���ֈړ����Ă���X���������܂����B8���̊ϑ����ʂ����(��II�͎Q��)�A�����p�ɂ�����A�T���c���̕��z�ɂ́A�k�Ε��ɂ���Ĉ����N�����ꂽ���x�t�����g�ɂ������@�\��A�~�Y�N���Q����і�����ɂ��ߐH����p���Ă��邱�Ƃ���������܂����B�܂��A�A�T���c���̕��z�ƕn�_�f����Ƃ̊W�͕s���ł����B10���̊ϑ��ł́A�~�Y�N���Q�������Ȃǂ̓��H���̃v�����N�g���͖w�Ǎ̏W����Ȃ��������Ƃ���A���̐����ɂ��ߐH�́A10���̃A�T���c���̕��z��̌Q���x�̌����ɂ���قǍ�p���Ă��Ȃ��ƍl�����܂��B�܂��A10���ɂ�����C��t�߂̕n�_�f����̋K�͂�8�����������Ȃ��Ƃ���A�c���̕��z�ƕn�_�f����Ƃ̊֘A���Ⴂ�Ǝv���܂��B�a�ʂɂ��Ă̓A�T���c���̐����̐����v���Ƃ͂Ȃ��Ă��Ȃ��ƍl�����邱�Ƃ���(��L4.1�Q��)�A�ϑ����Ԓ��A�a�s���ɂ��c���̑�ʎ��͋N���Ă��Ȃ��Ǝv���܂��B ��ʂɁA�����p�ɂ����Ėk�Ε����A�������ꍇ�ɂ́A���x���w�Ȑ�̊C���͘p�O�֗��o���A���x���w�Ȑ[�̊C���͗��o����₤���߂ɘp���֗������܂�(��2�`�p���ǁC1974�Q��)�B�ċG�̓����p�ł͓�Ε�����z���܂����A�k�Ε����A���������ɂ́A�����̍������w�̐����N�����錻�ۂ��A�p���������t����тɂ����Ă����Ίώ@����܂�(��2�`�p���ǁC1974�Q��)�B8���̊ϑ��ł��A�s���`�ՏF�ɉ������C��ɂ����āA�N���ɂ��Ǝv���錰���Ȗ��x�t�����g���ώ@����܂����i�}�h�h-2�j�B�ċG�ɖk�Ε����A�������Ƃ��Ă��A�A�T���c���̓t�����g�ɂ�镨���I�Ȏ����E���U�ɂ���ăt�����g���ӂ̊C��ɑؗ����A���ʂƂ��āA�p�O�֗��o���ɂ����Ȃ�ƍl�����܂��B10���̓����p�ł́A�k�Ε����A�����Ă������Ȗ��x�t�����g�͉͐쐅�̉e������\�w�ȊO�ł͖w�NJώ@����܂���ł���(�}III-2)�B����͐�������ԓI�Ɉ�l�ƂȂ�ɂƂ��Ȃ��A��t�̕ω����������Ȃ������Ƃɂ��Ǝv���܂�(�\III-1)�B�]���āA�t�����g�ɂ������E���U�A���Ȃ킿�A�A�T���c�����p���ɑؗ����錻�ۂ��N���ɂ����A��z����k�Ε��ɂ���Ė��x���w�Ȑ�ɂ���c���̑����͘p�O�֗��o���Ă���\��������܂��B10��23���̊ϑ��ł́A���x���w�͐��[8 m�t�߂Ɋώ@���ꂽ���Ƃ���(�}III-1)�A10��19������23���ɂ����Ċώ@���ꂽ���[0�`4 m�w�����4�`8 m�w�ɂ�����A�T���c���̕��z�́A�k�Ε��ɂ���Ĉ����N�����ꂽ��w���̘p�O�ւ̗��o�ɂ����̂Ɛ��@����܂��B�����āA8���Ɣ�r���ĕ��V���Ԃ������Ȃ邱�Ƃ�A�H�G�ɕp�ɂɗ��P����䕗�ɂ�镗�J���c���̘p�O�ւ̗��o���������Ă���Ǝv���܂��B�ΏƓI�ɁA���[8�`10
m�w�ɂ����Ċώ@���ꂽ�A�T���c���̕��z�͉��w���̗����ɂ����̂Ǝv���܂��B���x���w�Ȑ[�ɂ���A�T���c���ɂ��Ă͘p���ɗ��܂邱�Ƃ��ł���̂̊����������ƍl�����܂��B |
�T�D������
|
�O�Ԑ��ł͍ŋ�10�N�ȏ�ɓn���āA�A�T���̑��̐��ɑ���t�L�̏o�������ቺ���Ă��邱�Ƃ�����Ă��܂�(���H�C2002)�B�O�Ԑ��ł̓A�T���̎�c�������s���Ă��Ȃ����Ƃ���A�V�R�t�L�̏o�����̒ቺ�͓����p�ɂ�����c���̐��c�ߒ��̕ω������̂Ǝv���܂��B�����p�ł̓A�T���̎Y�����͏t�G�ƏH�G��2��ŁA��Y�����͏t�ƍl�����Ă��܂�(���H��C1993)�B�������A�~�G�ɂ�����A�T���̑�ʝˎ��ɂ���āA�����p�A���ɎO�Ԑ��ł͏t��̓A�T���̌����ʂ��Œ�ƂȂ鎞���ł��邱�Ƃ���A�Y�����͎�����A�H�G������1��ƂȂ��Ă���\��������܂�(���H�C2002�Q��)�B����ɁA�{�������ʂ������悤�ɁA�H�G�ɔ��������c���ɂ��ẮA�p���̐��⊱���Ȃǂɒ���ł���̂̊����͒Ⴂ�Ǝv���܂��B�����̂��Ƃ��t�L�̏o�����̒ቺ�A����ɂ́A�����p�ɂ�����A�T�������ʂ̌����̈���ƂȂ��Ă���\��������܂��B����A���l�v�Z�Ȃǂɂ��A���݂Ɖߋ��̗������A�T���c���̑ؗ��̏̈Ⴂ�Ȃǂ͂��邱�Ƃɂ��A���ڍׂȌ������ł���ł��傤�B |